-
1
المؤلفون: Mona Shastri, Ronak Hapani
المصدر: Radiology Case Reports
Radiology Case Reports, Vol 16, Iss 8, Pp 2053-2055 (2021)مصطلحات موضوعية: Deficiency syndrome, Ectrodactyly, Variable manifestations, business.industry, R895-920, Case Report, Tibial hemimelia, Anatomy, medicine.disease, 030218 nuclear medicine & medical imaging, body regions, 03 medical and health sciences, Medical physics. Medical radiology. Nuclear medicine, 0302 clinical medicine, Lobster claw deformity, Limb bones, Gollop-Wolfgang, Tibial aplasia/hemimelia, Rare case, Medicine, Radiology, Nuclear Medicine and imaging, Tibia, Ectrodactyly/lobster hand, business, 030217 neurology & neurosurgery
-
2Academic Journal
المؤلفون: Kantaputra, Piranit N., Carlson, Bruce M.
مصطلحات موضوعية: malformation of limb, oligodactyly, SHFM, lobster claw deformity, ectrodactyly, Genetics, Health Sciences
وصف الملف: application/pdf
Relation: Kantaputra, Piranit N.; Carlson, Bruce M. (2019). "Genetic regulatory pathways of split‐hand/foot malformation." Clinical Genetics 95(1): 132-139.; https://hdl.handle.net/2027.42/146859; Clinical Genetics; Lewandoski M, Sun X, Martin GR. Fgf8 signalling from the AER is essential for normal limb development. Nat Genet. 2000; 26: 460 ‐ 463.; Bell SM, Schreiner CM, Waclaw RR, Campbell K, Potter SS, Scott WJ. Sp8 is crucial for limb outgrowth and neuropore closure. Proc Natl Acad Sci U S A. 2003; 100: 12195 ‐ 12200.; Choi KS, Lee C, Maatouk DM, Harfe BD. Bmp2, Bmp4 and Bmp7 are co‐required in the mouse AER for normal digit patterning but not limb outgrowth. PLoS One. 2012; 7: e37826. https://doi.org/10.1371/journal.pone.0037826.; Schatz O, Langer E, Ben‐Arie N. Gene dosage of the transcription factor Fingerin (bHLHA9) affects digit development and links syndactyly to ectrodactyly. Hum Mol Genet. 2014; 23: 5394 ‐ 5401.; Kataoka K, Matsushima T, Ito Y, Sato T, Yokoyama S, Asahara H. Bhlha9 regulates apical ectodermal ridge formation during limb development. J Bone Miner Metab. 2018; 36: 64 ‐ 72.; Spielmann M, Kakar N, Tayebi N, et al. Exome sequencing and CRISPR/Cas genome editing identify mutations of ZAK as a cause of limb defects in humans and mice. Genome Res. 2016; 26: 183 ‐ 191.; Saito D, Yonei‐Tamura S, Kano K, Ide H, Tamura K. Specification and determination of limb identity: evidence for inhibitory regulation of Tbx gene expression. Development. 2002; 129: 211 ‐ 220.; Guerrini L, Costanzo A, Merlo GR. A symphony of regulations centered on p63 to control development of ectoderm‐derived structures. J Biomed Biotechnol. 2011; 864904: 1 ‐ 13. https://doi.org/10.1155/2011/864904.; Zeller R, López‐Ríos J, Zuniga A. Vertebrate limb bud development: moving towards integrative analysis of organogenesis. Nat Rev Genet. 2009; 10: 845 ‐ 858.; Gañan Y, Macias D, Basco RD, Merino R, Hurle JM. Morphological diversity of the avian foot is related with the pattern of msx gene expression in the developing autopod. Dev Biol. 1998; 196: 33 ‐ 41.; Kaltcheva MM, Anderson MJ, Harfe BD, Lewandoski M. BMPs are direct triggers of interdigital programmed cell death. Dev Biol. 2016; 411: 266 ‐ 276.; Khan A, Wang R, Han S, Ahmad W, Zhang X. A novel homozygous missense mutation in BHLHA9 causes mesoaxial synostotic syndactyly with phalangeal reduction in a Pakistani family. Hum Genome Var. 2017, 2017; 4: 17054. https://doi.org/10.1038/hgv.2017.54. eCollection.; Tickle C. Making digit patterns in the vertebrate limb. Nat Rev Mol Cell Biol. 2006; 7: 45 ‐ 53.; Hiscock TW, Tschopp P, Tabin CJ. On the formation of digits and joints during limb development. Devel Cell. 2017; 41: 459 ‐ 465.; Harfe BD, Scherz PJ, Nissim S, Tian H, McMahon AP, Tabin CJ. Evidence for an expansion‐based temporal Shh gradient in specifying vertebrate digit identities. Cell. 2004; 118: 517 ‐ 528.; Litingtung Y, Dahn RD, Li Y, Fallon JF, Chiang C. Shh and Gli3 are dispensable for limb skeleton formation but regulate digit number and identity. Nature. 2002; 418: 979 ‐ 983.; Conte D, Guerrini L, Merlo GR. Novel cellular and molecular interactions during limb development, revealed from studies on the split hand foot congenital malformation. In: Wu B, ed. New Discoveries in Embryology. INTECH; London, United Kingdom 2015: 125 ‐ 167.; Suzuki T, Hasso SM, Fallon JF. Unique SMAD 1/5/8 activity at the phalanx‐forming region determines digit identity. Proc Natl Acad Sci U S A. 2008; 105: 4185 ‐ 4190.; Harrison RG. On relations of symmetry in transplanted limbs. J Exp Zool. 1921; 32: 1 ‐ 136.; Swett FH. Determination of limb‐axes. Q Rev Biol. 1937; 2: 322 ‐ 339.; Long JE, Garel S, Depew MJ, Tobet S, Rubenstein JL. DLX5 regulates development of peripheral and central components of the olfactory system. J Neurosci. 2003; 23: 568 ‐ 578.; Ugur SA, Tolun A. Homozygous WNT10B mutation and complex inheritance in split‐hand/foot malformation. Hum Mol Genet. 2008; 17: 2644 ‐ 2653.; Lo Iacono N, Mantero S, Chiarelli A, et al. Regulation of Dlx5 and Dlx6 gene expression by p63 is involved in EEC and SHFM congenital limb defects. Development. 2008; 135: 1377 ‐ 1388.; Haro E, Delgado I, Junco M, et al. Sp6 and Sp8 transcription factors control AER formation and dorsal‐ventral patterning in limb development. PLoS Genet. 2014; 10: e1004468. https://doi.org/10.1371/journal.pgen.1004468.; Petit F, Sears KE, Ahituv N. Limb development: a paradigm of gene regulation. Nature Revs Genet. 2017; 18: 245 ‐ 258.; van Bokhoven H, Hamel BC, Bamshad M, et al. p63 gene mutations in EEC syndrome, limb‐mammary syndrome, and isolated split hand‐split foot malformation suggest a genotype‐phenotype correlation. Am J Hum Genet. 2001; 69: 481 ‐ 492.; Gieruszczak‐Bialek D, Oldak M, Skorka A, Waligora J, Korniszewski L. Fibular aplasia with ectrodactyly‐broadening the clinical spectrum. Eur J Med Genet. 2006; 49: 83 ‐ 86.; Shamseldin HE, Faden MA, Alashram W, Alkuraya FS. Identification of a novel DLX5 mutation in a family with autosomal recessive split hand and foot malformation. J Med Genet. 2012; 49: 16 ‐ 20.; Wang X, Xin Q, Li L, et al. Exome sequencing reveals a heterozygous DLX5 mutation in a Chinese family with autosomal‐dominant split‐hand/foot malformation. Eur J Hum Genet. 2014; 22: 1105 ‐ 1110.; Ullah A, Hammid A, Umair M, Ahmad W. A novel heterozygous intragenic sequence variant in DLX6 probably underlies first case of autosomal dominant split‐hand/foot malformation type 1. Mol Syndromol. 2017; 8: 79 ‐ 84.; Villanueva C, Jacobson‐Dickman E, Xu C, et al. Congenital hypogonadotropic hypogonadism with split hand/foot malformation: a clinical entity with a high frequency of FGFR1 mutations. Genet Med. 2015; 17: 651 ‐ 659.; Klopocki E, Lohan S, Doelken SC, et al. Duplications of BHLHA9 are associated with ectrodactyly and tibia hemimelia inherited in non‐Mendelian fashion. J Med Genet. 2012; 49: 119 ‐ 125.; Malik S, Percin FE, Bornholdt D, et al. Mutations affecting the BHLHA9 DNA‐binding domain cause MSSD, mesoaxial synostotic syndactyly with phalangeal reduction, Malik‐Percin type. Am J Hum Genet. 2014; 95: 649 ‐ 659.; Huh JK, Chung MS, Baek GH, Oh JH, Lee YH, Gong HS. Cleft hand in Kabuki make‐up syndrome: case report. J Hand Surg Am. 2011; 36: 653 ‐ 657.; Conte D, Garaffo G, Lo Iacono N, et al. The apical ectodermal ridge of the mouse model of ectrodactyly Dlx5;Dlx6−/− shows altered stratification and cell polarity, which are restored by exogenous Wnt5a ligand. Hum Mol Genet. 2016; 25: 740 ‐ 754.; Umair M, Ullah A, Abbas S, Ahmad F, Basit S, Ahmad W. First direct evidence of involvement of a homozygous loss‐of‐function variant in the EPS15L1 gene underlying split‐hand/split‐foot malformation. Clin Genet. 2018; 93: 699 ‐ 702.; Bakkers J, Hild M, Kramer C, Furutani‐Seiki M, Hammerschmidt M. Zebrafish DeltaNp63 is a direct target of Bmp signalling and encodes a transcriptional repressor blocking neural specification in the ventral ectoderm. Dev Cell. 2002; 2: 617 ‐ 627.; Mills AA, Zheng B, Wang XJ, Vogel H, Roop DR, Bradley A. p63 is a p53 homologue required for limb and epidermal morphogenesis. Nature. 1999; 398: 708 ‐ 713.; Yang A, Schweitzer R, Sun D, et al. p63 is essential for regenerative proliferation in limb, craniofacial and epithelial development. Nature. 1999; 398: 714 ‐ 718.; Kjaer KW, Hansen L, Schwabe GC, et al. Distinct CDH3 mutations cause ectodermal dysplasia, ectrodactyly, macular dystrophy (EEM syndrome). J Med Genet. 2005; 42: 292 ‐ 298.; Ullah A, Ullah MF, Khalid ZM, Ahmad W. Novel heterozygous frameshift mutation in distal‐less homeobox 5 underlies isolated split hand/foot malformation type 1. Pediatr Int. 2016; 58: 1348 ‐ 1350.; Merlo GR, Paleari L, Mantero S, et al. Mouse model of split hand/foot malformation type I. Genesis. 2002; 33: 97 ‐ 101.; Robledo RF, Rajan L, Li X, Lufkin T. The Dlx5 and Dlx6 homeobox genes are essential for craniofacial, axial, and appendicular skeletal development. Genes Dev. 2002; 16: 1089 ‐ 1101.; Panganiban G, Rubenstein JL. Developmental functions of the distal‐less/dlx homeobox genes. Development. 2002; 129: 4371 ‐ 4386.; Restelli M, Lopardo T, Lo Iacono N, et al. DLX5, FGF8 and the Pin1 isomerase control ΔNp63α protein stability during limb development: a regulatory loop at the basis of the SHFM and EEC congenital malformations. Hum Mol Genet. 2014; 23: 3830 ‐ 3842.; Restelli M, Molinari E, Marinari B, et al. FGF8, c‐Abl and p300 participate in a pathway that controls stability and function of the ΔNp63α protein. Hum Mol Genet. 2015; 24: 4185 ‐ 4197.; Crackower MA, Motoyama J, Tsui LC. Defect in the maintenance of the apical ectodermal ridge in the Dactylaplasia mouse. Dev Biol. 1998; 201: 78 ‐ 89.; Paina S, Garzotto D, DeMarchis S, et al. Wnt5a is a transcriptional target of dlx homeogenes and promotes differentiation of interneuron progenitors in vitro and in vivo. J Neurosci. 2011; 31: 2675 ‐ 2687.; Geetha‐Loganathan P, Nimmagadda S, Scaal M. Wnt signaling in limb organogenesis. Organogenesis. 2008; 4: 109 ‐ 115.; Barrow JR, Thomas KR, Boussadia‐Zahui O, et al. Ectodermal Wnt3/beta‐catenin signalling is required for the establishment and maintenance of the apical ectodermal ridge. Genes Dev. 2003; 17: 394 ‐ 409.; Kawakami Y, Capdevila J, Büscher D, Itoh T, Rodríguez Esteban C, Izpisúa Belmonte JC. WNT signals control FGF‐dependent limb initiation and AER induction in the chick embryo. Cell. 2001; 104: 891 ‐ 900.; Kantaputra PN, Hutsadaloi A, Kaewgahya M, et al. WNT10B mutations associated with isolated dental anomalies. Clin Genet. 2018; 93: 992 ‐ 999.; Witte F, Dokas J, Neuendorf F, Mundlos S, Stricker S. Comprehensive expression analysis of all Wnt genes and their major secreted antagonists during mouse limb development and cartilage differentiation. Gene Expr Patterns. 2009; 9: 215 ‐ 223.; Gros J, Hu JK‐H, Vinegoni C, Feruglio PF, Weissleder R, Tabin CJ. WNT5A/JNK and FGF/MAPK pathways regulate the cellular events shaping the vertebrate limb bud. Curr Biol. 2010; 20: 1993 ‐ 2002.; Ho HY, Susman MW, Bikoff JB, et al. Wnt5a‐Ror‐Dishevelled signalling constitutes a core developmental pathway that controls tissue morphogenesis. Proc Natl Acad Sci U S A. 2012; 109: 4044 ‐ 4051.; Yamaguchi TP, Bradley A, McMahon AP, Jones S. A. Wnt5a pathway underlies outgrowth of multiple structures in the vertebrate embryo. Development. 1999; 126: 1211 ‐ 1223.; Kengaku M, Capdevila J, Rodriguez‐Esteban C, et al. Distinct WNT pathways regulating AER formation and dorsoventral polarity in the chick limb bud. Science. 1998; 280: 1274 ‐ 1277.; Talamillo A, Delgado I, Nakamura T, et al. Role of Epiprofin, a zinc‐finger transcription factor, in limb development. Dev Biol. 2010; 337: 363 ‐ 374.
-
3
المؤلفون: V. Rami Reddy, B.K. Chandrasekhar Reddy
المصدر: International Journal of Human Genetics. 4:219-226
مصطلحات موضوعية: Genetics, Polydactyly, Genetic traits, Pedigree chart, Biology, medicine.disease, Penetrance, Variable Expression, Lobster claw deformity, medicine, Anonychia, Left forearm, Genetics (clinical)
-
4
المؤلفون: Sahana M Srinivas, Ravi Hiremagalore
المصدر: Indian Journal of Dermatology, Vol 60, Iss 1, Pp 106-106 (2015)
Indian Journal of Dermatologyمصطلحات موضوعية: lobster-claw hand, integumentary system, Skeletal anomalies, business.industry, Dermatology, Anatomy, lcsh:RL1-803, medicine.disease, Focal dermal hypoplasia, Lobster claw hand, Umbilical hernia, Lobster claw deformity, Hypoplastic nails, Rare case, lcsh:Dermatology, medicine, Goltz syndrome, Syndactyly, business, E-IJD Case Report
-
5
المؤلفون: Ashish Agrawal, Seema Agrawal, Rajat Singh, Rahul Agrawal, Romi Agrawal
المصدر: Indian Journal of Dental Research, Vol 25, Iss 2, Pp 243-247 (2014)
مصطلحات موضوعية: Adult, Foot Deformities, Male, Ectodermal dysplasia, animal structures, Ectrodactyly, Biology, Split-Hand/Foot Malformation, Rare case, medicine, Humans, Abnormalities, Multiple, endogenous erythroid colony syndrome, Lobster claw, General Dentistry, p63, fungi, food and beverages, Syndrome, General Medicine, Anatomy, Hand Deformities, split hand foot malformation, medicine.disease, lcsh:RK1-715, stomatognathic diseases, Lobster claw deformity, lcsh:Dentistry, lobster claw
-
6
المؤلفون: Luella Nadelhoffer, Edith L. Potter
المصدر: Journal of Heredity. 38:331-335
مصطلحات موضوعية: Lobster claw deformity, Genetics, Anatomy, Biology, Molecular Biology, Genetics (clinical), Biotechnology
-
7
المؤلفون: Frank A Walker, Otto A Wiegmann
المصدر: Journal of Pediatric Ophthalmology & Strabismus. 7:79-85
مصطلحات موضوعية: Ophthalmology, Lobster claw deformity, Nasolacrimal Obstruction, business.industry, Pediatrics, Perinatology and Child Health, Medicine, General Medicine, Anatomy, business
-
8
المؤلفون: Sharpe Ld
المصدر: Journal of The Royal Naval Medical Service. 44:59-61
مصطلحات موضوعية: Lobster claw deformity, business.industry, Medicine, General Medicine, Anatomy, business
-
9
المؤلفون: S. L. Pugsley
المصدر: Laboratory animals. 18(4)
مصطلحات موضوعية: General Veterinary, Monkey Diseases, Limb Deformities, Congenital, Extremities, Anatomy, Biology, biology.organism_classification, Radiography, Lobster claw deformity, Pregnancy, Callitrichinae, London, Silvery marmoset, Animals, Animal Science and Zoology, Animals, Zoo, Female, Fetal Death, Callithrix argentata
-
10
-
11
-
12
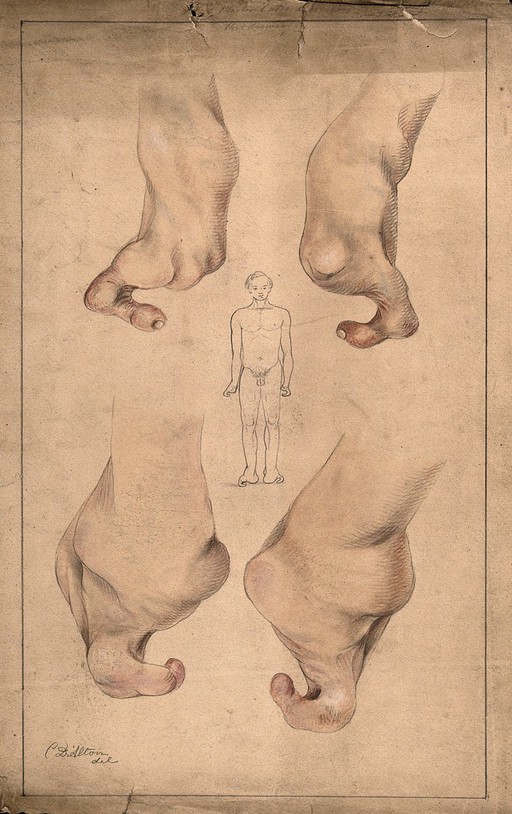
المؤلفون: Christopher D'Alton
المصدر: V0009996
-
13Image
المؤلفون: Karl Pearson
المصدر: L0033874
مصطلحات موضوعية: Genetic disease, Inheritance, Lobster claw deformity, Eugenics, Wills
Relation: http://wellcomeimages.org/ixbin/hixclient.exe?MIROPAC=L0033874; L0033874; External Reference; http://catalogue.wellcomelibrary.org/record=b1036328
